W karbonie z najprymitywniejszych owodniowców niezależnie powstały synapsydy, w których czaszce występował jeden (dolny) otwór skroniowy za każdym okiem, oraz zauropsydy. Synapsydy były dominującą grupą kręgowców lądowych w okresie permu i na początku triasu; w erze mezozoicznej dały początek ssakom.
Zauropsydy rozdzieliły się na dwie linie ewolucyjne: anapsydy, charakteryzujące się czaszką pozbawioną otworów skroniowych, oraz diapsydy. Wśród anapsydów pojawiły się m.in.: parejazaury, wymarłe na przełomie permu i triasu, oraz nadrodzina Procolophonoidea wymarła pod koniec triasu. Z kolei diapsydy wykształciły czaszkę z dwoma parami otworów skroniowych (pary: dolna i górna). Dzięki obecności otworów skroniowych czaszka była lżejsza, a mięśnie miały więcej miejsca; w rezultacie szczęka stawała się silniejsza i bardziej ruchliwa. Diapsydy rozdzieliły się na kilka linii ewolucyjnych, w tym m.in. grupy Younginiformes i Ichthyopterygia (choć Merck, 1997 uważa Ichthyopterygia za bazalne archozauromorfy, zaś z analiz kladystycznych przeprowadzonych przez Maischa, 2010 wynika, że Ichthyopterygia mogły należeć do anapsydów) oraz klad Sauria, obejmujący dwie główne linie:
Przedmiotem sporów pozostaje pozycja filogenetyczna żółwi. Tradycyjnie były one zaliczane do anapsydów i uznawane za potomków Procolophonoidea lub parejazaurów albo za takson siostrzany do eunotozaura. Niektóre analizy filogenetyczne sugerują jednak, że w rzeczywistości są one diapsydami należącymi do kladu Sauria. Analizy wykorzystujące cechy morfologiczne wskazują na przynależność żółwi do lepidozauromorfów, generalnie sugerując bliskie pokrewieństwo z zauropterygami (choć w analizie Müllera, 2004 żółwie są bliżej spokrewnione z lepidozaurami niż z zauropterygami); natomiast analizy opierające się na badaniach genetycznych sugerują przynależność źółwi do archozauromorfów.
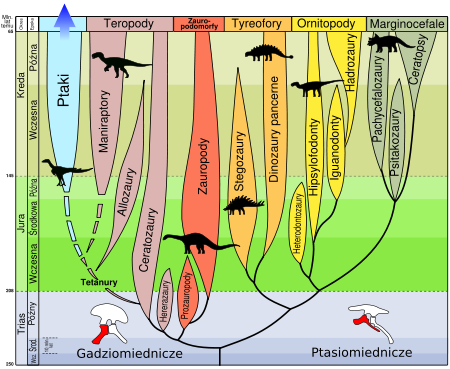
Kladogram ilustrujący uproszczone „drzewo rodowe” gadów według Laurina i Gauthiera (1996), za Tree of Life Web Project: